
Abstract
There is a significant global upsurge in the number and proportion of older persons in the population. With this comes an increasing prevalence of age-related conditions which pose a major challenge to healthcare systems. The development of anti-ageing treatments may help meet this challenge by targeting the ageing process which is a common denominator to many health problems. Cannabis-like compounds (cannabinoids) are reported to improve quality of life and general well-being in human trials, and there is increasing preclinical research highlighting that they have anti-ageing activity. Moreover, preclinical evidence suggests that endogenous cannabinoids regulate ageing processes. Here, we review the anti-ageing effects of the cannabinoids in various model systems, including the most extensively studied nematode model, Caenorhabditis elegans. These studies highlight that the cannabinoids lengthen healthspan and lifespan, with emerging evidence that they may also hinder the development of cellular senescence. The non-psychoactive cannabinoid cannabidiol (CBD) shows particular promise, with mechanistic studies demonstrating it may work through autophagy induction and activation of antioxidative systems. Furthermore, CBD improves healthspan parameters such as diminishing age-related behavioural dysfunction in models of both healthy and accelerated ageing. Translation into mammalian systems provides an important next step. Moreover, looking beyond CBD, future studies could probe the multitude of other cannabis constituents for their anti-ageing activity.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ageing is a complex biological process that involves the gradual deterioration of cellular and molecular processes over time. This process is influenced by a variety of factors, including genetics, lifestyle and environmental factors [1]. During ageing, our bodies gradually become less efficient at repairing damage and defending against disease [2], which can result in an overall decline in health and an increased risk of chronic conditions such as cardiovascular disease [3], diabetes [4] and cancer [5]. Age-related changes can affect almost every system in the body, including the immune system [6], the cardiovascular system [7] and the nervous system [8]. These changes can result in a variety of functional impairments, such as decreased mobility, impaired cognition and reduced sensory function, that can significantly impact the quality of life and independence [9, 10].
As the global population continues to age, there is increasing interest in identifying strategies that can promote healthy ageing and extend lifespan. Because of the challenges brought by an ageing society, the United Nations declared 2021–2030 the UN Decade in Healthy Ageing and has called for society to increase efforts to develop innovative means to improve both the healthspan and lifespan of the human race. Basic research can contribute to this effort via the discovery of chemical agents that improve healthy ageing. The development of anti-ageing therapies provides a paradigm shift in treating disease. As many health conditions are age-related, diminishing age-related decline might provide a common means to prevent a spectrum of diseases and comorbidities that plague the community.
The legalisation of medicinal cannabis around the world has prompted the question of whether cannabinoids (cannabis-like drugs) might promote longevity and mitigate age-related decline. There is an emerging picture from clinical trials on medicinal cannabis products that they improve the general quality of life and well-being of patients [11]. However, controlled studies are needed to specifically address whether cannabinoids can extend the healthspan (the number of years having functional health) and lifespan. In this review, we explore the potential benefits of cannabinoids to healthy ageing and ageing-related disease by examining whether the endogenous cannabinoid (endocannabinoid) system and exogenous cannabinoids extend healthspan and lifespan in preclinical models. While beyond the scope of the present review, it is important to acknowledge that cannabinoid exposure may negatively impact early development. The interested reader is encouraged to see the following excellent reviews on the developmental impacts and toxicity of cannabinoid exposure at different ages (e.g. prenatal, adolescent or adult) [12,13,14,15].
Background on the cannabinoids
Cannabinoids are chemical compounds that are found in the Cannabis plant [16]. Over 100 plant cannabinoids (phytocannabinoids) have been elucidated, with the most characterised being the main psychoactive constituent, Δ9-tetrahydrocannabinol (THC), and the non-intoxicating constituent cannabidiol (CBD). Whilst being structural isomers, these major cannabinoids have a distinct pharmacology and potential therapeutic applications [17,18,19]. At present, THC-containing products are registered treatments for chemotherapy-induced nausea and vomiting, and spasticity associated with multiple sclerosis, whereas CBD is a registered drug to treat intractable epilepsies such as Dravet syndrome and Lennox-Gastaut syndrome [20, 21]. There is currently intensive research activity exploring additional therapeutic applications for the cannabinoids. These include studies on molecules such as cannabidiolic acid, cannabigerolic acid, cannabigerol (CBG), cannabichromene (CBC) and cannabinol (CBN), among others [22,23,24,25,26].
One of the main modes of action of the phytocannabinoids THC and CBD is to modulate the body’s endocannabinoid system (ECS) (Fig. 1) [27]. This is a complex signalling system that plays a role in regulating various physiological processes such as mood, appetite, pain sensation, sleep and immune function [28,29,30,31]. The ECS is composed of a network of receptors, endocannabinoids and enzymes that work together to maintain homeostasis. The two primary receptors of the ECS are cannabinoid receptor type-1 (CB1R) and cannabinoid receptor type-2 (CB2R), which are found throughout the body, including in the brain, immune system and peripheral tissues [32, 33]. Two of the most extensively studied endogenous ligands are the endocannabinoids anandamide (AEA) and 2-arachidonoylglycerol (2-AG), as well as the enzymes responsible for the metabolism of endocannabinoids, including fatty acid amide hydrolase (FAAH) and monoacylglycerol lipase (MAGL). THC directly activates cannabinoid receptors CB1R and CB2R to produce its pharmacological actions, whereas CBD is thought to be an indirect agonist that boosts endogenous concentrations of the endocannabinoids like AEA via inhibition of fatty acid-binding proteins (FABPs) [34], such as FABP5 and FABP7 [35]. FABP5 plays a key role in retrograde endocannabinoid signaling and acts as a carrier of endocannabinoids at synapses in the central nervous system [36]. It should also be recognised that CBD has a multitude of pharmacological actions beyond the ECS, including antagonism of G protein-coupled receptor 55 (GPR55), activation of transient receptor potential vanilloid-1 (TRPV1) channels and positive allosteric modulation of γ-aminobutyric acid sub-type A (GABAA) receptors [37,38,39,40,41,42].

Chemical structures of endocannabinoids (produced naturally by the body) and major phytocannabinoids (produced by plants). THC, Δ9-tetrahydrocannabinol; CBD, cannabidiol; CBN, cannabinol; CBG, cannabigerol; CBC, cannabichromene
Ageing and the ECS
Several changes occur in the ECS during ageing that may have health implications. In the human brain, CB1R mRNA expression gradually decreased with age in the prefrontal cortex (PFC) (Fig. 2) [43]. In addition, aged rats displayed increased CB1R protein expression in the entorhinal and temporal cortices but decreased expression in the postrhinal cortex compared to young rats [44]. Further, aged female mice showed increased CB1R mRNA expression in subregions of the cortex compared to young mice [45]. However, aged mice exhibited decreased CB1R mRNA expression in the lateral septal area compared to younger animals [45].

Ageing decreased mRNA expression of cannabinoid CB1R but not CB2R in the PFC of humans with ages ranging from birth to 50 years. PFC, the prefrontal cortex; CB1R, cannabinoid CB1 receptors; CB2R, cannabinoid CB2 receptors; n.s., not significant
Age-related changes in the brain ECS are accompanied by decrements in physiological function. Genetic deletion of CB1R in mice led to an earlier decline in cognitive and memory function specific to both reward and aversion-driven learning [46,47,48]. Young mice lacking CB1R exhibited comparable or even improved learning and memory performance compared to age-matched wild-type (WT) mice in various learning and memory tasks. In contrast, mature mice lacking CB1R performed significantly worse than age-matched WT mice in the same tasks. Additionally, the rapid decline in cognitive functions observed in mature CB1R-deficient mice was accompanied by a loss of neurons in the CA1 and CA3 regions of the hippocampus [46]. A noteworthy finding suggests that a decrease in CB1 signaling specifically in the limbic forebrain contributed to age-related decline in appetite for food and alcohol [49].
In addition to the involvement of CB1R signaling in the ageing process, CB2R may also play a role. CB2R is abundantly expressed in immune cells, including the brain’s immune cells, the microglia. CB2R is upregulated during inflammation and could contribute to inflammaging, the chronic, low-grade inflammation that occurs during ageing. Consistent with this notion, unlike WT mice that display an age-related decline in social memory, CB2R knockout mice were resistant to such a decline and even showed some improvement [50]. This effect was accompanied by various molecular and morphological changes in the microglia of the CB2R knockout mice. Interestingly, another study revealed that β-caryophyllene, a natural agonist of CB2R, reversed age-related impairments in working memory and reduced circulating inflammatory cytokine concentrations in aged mice [51].
It is uncertain whether tissue concentrations of the endocannabinoids are altered during the ageing process. Concentrations of AEA and 2-AG did not exhibit age-dependent changes in various brain regions such as the hypothalamus, limbic forebrain, amygdala and cerebellum [49]. A recent study investigated changes in the ECS during ageing in the mouse brain [47]. The ECS was most impacted in middle-aged mice compared to young adult mice. Specifically, middle-aged mice displayed a prominent decrease in 2-AG in the hippocampus, whereas AEA was decreased in other regions. There is evidence that ageing influences the expression and activity of enzymes that synthesize and degrade endocannabinoids. For example, aged animals exhibited reduced expression of the 2-AG synthesising enzyme DAGL [47], but increased expression of the AEA synthesising enzyme N-acyl phosphatidyl ethanolamide-phospholipase D (NAPE-PLD) compared to younger mice [52]. Another study reported age-related alterations in ECS components in female reproductive organs [53]. In conclusion, the ECS undergoes various changes during the ageing process and appears functionally implicated in the protection against age-related physiological decline.
Model systems for studying ageing
Various model systems, such as fruit fly (Drosophila) [54, 55], nematodes (Caenorhabditis elegans) [56] and zebrafish (Danio rerio) [57], have offered valuable insights into the ageing process. These model organisms are particularly advantageous due to their relatively short lifespans, which have enabled the elucidation of the molecular, genetic and physiological factors that influence ageing and age-related diseases [58,59,60]. Rodents are also employed as an ageing model because of their genetic similarity to humans, and genetic rodent models of accelerated ageing have been developed. However, while important, rodent ageing studies are time-consuming and of much lower throughput in the discovery phase of ageing research [61]. Lifespan is the number of years someone lives from birth until death, while healthspan refers to the period of life spent in good health, free from age-related diseases and disabilities [62]. Model systems used to study ageing incorporate both measurements of lifespan and healthspan, as there is no point in increasing longevity without a corresponding improvement in overall health and well-being.
Here, we provide a more detailed background on the nematode C. elegans, which has been the most widely used model when studying the effects of cannabinoids on lifespan and healthspan. More generally, the C.elegans model is used in ageing research due to the ability to rapidly discern age-related physiological decline over their short lifespan (~ 3 weeks). C. elegans is a free-living nematode with a small size (1 mm in length) and is easy to maintain in laboratory settings. C. elegans can develop from eggs to adults in about 3 days. The growth of C. elegans contains four larval stages (L1–L4) and an adult stage (Fig. 3). These convenient features make it particularly suitable as a model system for studies on ageing, age-related diseases and mechanisms of longevity.

The life cycle of C. elegans at 20 °C and age-related changes that are measured in geroscience research. Under normal laboratory conditions, an adult hermaphrodite produces roughly 300 eggs that hatch after 14 h. In favourable conditions, C. elegans goes through four larval stages (L1 to L4) to the adult in 3–5 days. In unfavourable conditions (overcrowding/low food/high temperature), the L1 larva may choose the dauer larva, which can live for several months. The overall lifespan of C. elegans is 2–3 weeks. Age-related changes in the nervous system, muscle and reproductive system
Age-related changes are evident in various tissues of C. elegans, including the cuticle (skin) and the reproductive and nervous systems [63] (Fig. 3). The ageing intestine of C. elegans displays deterioration of intestinal microvilli [64]. Fertility significantly declines with age, and the reproductive system deteriorates due to factors such as sperm shortage and reduced oocyte size and quality [65, 66]. During the ageing process, unmated C. elegans experience a decline in brood size [67, 68]. Age-related morphological changes in neurons have been extensively documented in C. elegans. Although earlier reports suggested no neuron loss or axon deficits in ageing worms, recent studies have identified delicate yet consistent age-dependent alterations in the organism’s neurons [69]. In adult hermaphrodites, dopaminergic and touch receptor neurons, exhibit age-related changes such as gradual beading, blebbing and branching [70]. Thus, using C. elegans models provides a clear benefit, as it allows for the study of age-related neurodegeneration and the discovery of potential therapeutic compounds for ageing and age-related diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) (Fig. 4).

Neurodegenerative disease models in C. elegans used to study cannabinoid effects on healthspan and lifespan. Created with www.biorender.com
Phenotypic testing in nematodes has led to the discovery of anti-ageing compounds, and some of these compounds are already in clinical use. For example, metformin, an anti-diabetic drug, extends lifespan and healthspan in nematodes and in mouse models of ageing [71, 72]. Beyond its anti-diabetic effects, population studies have found that metformin use decreased the risk of developing cancer [73], cognitive impairment and dementia [74] and cardiovascular disease [75]. Metformin has also been reported to reduce the risk of spontaneous abortions and obstetric complications [72]. There is increasing evidence that metformin improves general well-being beyond its diabetic mode of action and through classic anti-ageing pathways [72]. This has prompted calls for evidence on whether metformin has health benefits in non-diabetic patients. In one smaller scale, double-blind placebo-controlled study in healthy elderly participants, metformin was shown to impact non-metabolic pathways linked to ageing [76]. Providing a roadmap for future clinical studies, a larger scale, multi-site trial called TAME (targeting ageing with metformin) aims to enrol 3000 people in the 65–79 age range with a primary endpoint of time until age-related morbidity (e.g. coronary heart disease, cancer, dementia) and mortality [72]. Three other trials (phase 1 and 2) are also underway looking at whether metformin improves physical performance and reduces frailty [72].
Anti-ageing effects of endogenous cannabinoids in C. elegans
C.elegans contain an ancestral endocannabinoid system including the endocannabinoid signalling lipids AEA and 2-AG [77]. The AEA biosynthetic enzyme NAPE-PLD [78] has two functional orthologs in C. elegans, namely nape-1 and nape-2, which have overlapping expression in the pharynx and brain interneurons [79]. Overexpression of nape-1 and nape-2 in worms produced distinct growth phenotypes at different ambient temperatures [80]. Overexpression of nape-1 reduced lifespan only at 25 °C, whereas overexpression of nape-2 extended lifespan at 15 °C.
The nematodes have an ortholog of the AEA catabolic enzyme FAAH, known as faah-1. Faah-1 is predominantly expressed in the pharynx where it is co-expressed with nape-2 [79]. Reducing the levels of faah-1 using RNA interference (RNAi) in worms increased concentrations of N-acyl ethanolamines (NAEs), including AEA. Dietary restriction, known to diminish NAE concentrations, extended the lifespan of worms [81, 82]. Further, the overexpression of faah-1 in the pharynx also increased lifespan [79]. The ortholog of the 2-AG biosynthetic enzymes DAGLα/DAGLβ has an orthologue in C. elegans named dagl-1. Overexpression of dagl-1 in nematodes increased lifespan, whereas the genetic downregulation of dagl-1 decreased lifespan and resistance to paraquat-induced oxidative stress [83]. It is worth noting that C. elegans lacks an ortholog for the 2-AG-metabolising enzyme MAGL. Interestingly, a recent study reported that the MAGL inhibitor JZL184 extended the lifespan of C. elegans [84]. Notably, activity-based protein profiling determined faah-4 (ortholog of FAAH in C. elegans) as the major target of JZL184 in this study. Collectively, these findings indicate that C. elegans may have a complete and functional endocannabinoid signalling pathway, similar to mammals, which regulates ageing and lifespan.
The ECS in C. elegans also subserves locomotion, nociception, reproduction, feeding and axon regeneration. AEA was reported to bind the G-protein-coupled receptors (GPCRs) NPR-19 and NPR-32 in C. elegans, and the activation of these receptors suppressed axon regeneration [85]. Although the nematode NPR-19 ortholog has only 23% similarity to human CB1R, an analysis revealed that the crucial amino acids responsible for AEA binding seem to be preserved across species [86, 87]. Furthermore, 2-AG or AEA activated NPR-19 to inhibit nociception and feeding. The endocannabinoids also work through other receptors such as OCTR-1 (α2A-adrenergic-like octopamine receptor) and SER-4 (5-HT1A-like serotonin receptor) to influence nociception and locomotion [87]. Studies found that 2-AG acted as an endogenous modulator of TRPV signal transduction, influencing sterol mobilisation via the IGF-1 signalling pathway [88] and stimulating cholesterol trafficking [89] in nematodes. AEA has a bidirectional effect on feeding in C. elegans, which is mediated by NPR-19 [90]. Interestingly, the effect of genetic deletion of NPR-19 on food preference was rescued by either introducing overexpression of NPR-19 or overexpression of the human CB1 receptor. This reinforces that there is a high degree of conservation of nematode and human endocannabinoid systems. Furthermore, functional conservation of the ECS for regulating food preferences was linked to the modulation of AWC chemosensory neurons [90].
Exogenous plant cannabinoid effects on lifespan in C. elegans
There is accumulating research showing that plant cannabinoids extend lifespan and healthspan in nematodes. Much of the research has focussed on the non-psychoactive cannabinoid constituent CBD, which is being sold as an over-the-counter wellness product in many countries [18, 91]. Recent evidence shows that CBD increased longevity in C.elegans in both wild-type (“healthy”) nematodes and also in nematode models of neurodegeneration (Table 1). We will now review these studies in greater detail by examining the magnitude of the effect of CBD in C.elegans models, as well as examining potential anti-ageing mechanisms in this species.
The first lifelong “toxicity” study of CBD in N2 (Bristol wild-type strain) C. elegans was published in 2021 [92]. Whole-life treatment with CBD (10–100 µM) increased lifespan, and a maximum lifespan extension of 18% was observed at 40 µM. CBD also increased thermotolerance as it protected against temperature-induced mortality. Another study in the N2 strain showed 100 µM CBD increased survival at various time points of daily exposure [93]. In addition, CBD also extended lifespan extension at lower concentrations (1 and 5 µM) in the N2 strain [94]. Overall, these results are very promising when you consider that metformin increased mean lifespan by 40% at a much higher concentration (50 mM) in nematodes [71, 72]. Results are also comparable to thioflavin T, which is another very promising anti-ageing compound that has robust lifespan-extending effects in C. elegans (25% increase in lifespan at 50 µM) [99]. The ability of CBD to extend the lifespan in C.elegans has now been replicated across 3 independent laboratories. Moreover, the effects of CBD were robust across different developmental treatment regimens; positive studies have administered CBD from the egg stage, but also from early adulthood [92,93,94].
Ageing is the leading risk factor for neurodegenerative diseases, such as AD and PD [100], and the anti-ageing effects of CBD have been observed in worm models of neurodegeneration (Table 1 and Fig. 4). CBD 5 µM increased the lifespan in both the control strain and in an amyloid β (Aβ) overexpression strain of AD by 40% and 25.6% respectively [95]. Dopaminergic neuron degeneration is often observed in AD, and is a contributing factor to cognitive impairment during the early stages of the disease [101, 102]. It was shown that CBD 5 µM reduced Aβ-induced degeneration of dopaminergic neurons [95].
To study PD in C. elegans, neurotoxin 6-hydroxydopamine (6-OHDA) models are used which model the selective degeneration of dopaminergic neurons observed in PD patients. CBD at 25, 50 and 100 µM increased lifespan by 18.3%, 32.7% and 45.1% respectively in a concentration-dependent manner in the 6-OHDA dopamine-depletion model of PD [96]. A similar lifespan extension was reported for CBD in the BZ555 strain treated with 6-OHDA (the BZ555 strain is used to visualise dopamine neurons with a GFP tag on the dat-1 promoter) [96]. CBD partially recovered the loss of dopaminergic neurons in 6-OHDA-treated BZ555 worms, as well as food-sensing behaviour deficits [96]. Cannabidivarin (CBDV), is a structural analog of CBD, and showed longevity benefits in the α-synuclein aggregation PD model of C. elegans at 10 and 50 µM concentrations [98]. Moreover, CBDV reduced 6-OHDA-induced reductions in the soma size of dopamine neurons and deficits in body bends’ behaviour in BZ555 nematodes [98].
Mechanisms for the anti-ageing effects of cannabinoids on lifespan in C. elegans
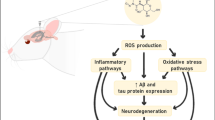
Anti-ageing mechanisms of CBD have been elucidated, with nematode research showing that CBD induces autophagy and reduces oxidative stress (Table 1, Fig. 5). Autophagy is a cellular process that removes impaired cellular organelles and debris. Impaired autophagy promotes ageing, whereas increased autophagy has anti-ageing effects [103,104,105]. Most autophagy genes have orthologs in worms as the process is highly conserved across species [106]. Studies have examined the effects of autophagy genes on CBD-induced lifespan extension using RNAi to knockdown specific genes. Conditional knockdown of bec-1, vps-34 and sqst-1 abolished CBD-induced lifespan extension [94]. It is noteworthy that these genes play a pivotal role in the intricate process of autophagy, influencing the formation of autophagic vesicles and the overall functionality of the autophagic machinery [107]. CBD also promoted longevity through the sir-2.1-dependent induction of autophagy (sir-2.1 is an ortholog of sirtuin 1 (SIRT1) in C. elegans). SIRT1 regulates autophagy via modulation of the mammalian target of rapamycin (mTOR) and AMP-activated protein kinase (AMPK) activity [94].

A summary of the known anti-ageing mechanisms of CBD in C. elegans. Created with www.biorender.com
Reactive oxygen species (ROS), including hydrogen peroxide (H2O2), hydroxyl radical (HO) and superoxide anions, can cause damage to cellular components, including DNA, proteins and lipids [108]. This damage can accumulate over time and contribute to the ageing process [109]. Several studies using C.elegans have demonstrated that the antioxidant properties of CBD contribute to its pro-longevity effects. One study investigated the glyoxalase pathway, an intrinsic antioxidant system, in the anti-ageing effects of CBD in worms [93]. The glyoxalase pathway involves glyoxalase enzymes which metabolise methylglyoxal (MG), a toxic metabolite that accumulates with age and may contribute to neurodegeneration [110]. MG treatment reduced the lifespan of various C. elegans strains, whereas treatment with 100 µM CBD attenuated the adverse effects of MG on lifespan. This finding was attributed to CBD enhancing the glyoxalase pathway via increasing the expression of glyoxalase enzymes. It was also found that CBD reduced the MG-induced increase in Aβ expression and oxidative stress in C. elegans.
CBD reduced ROS in N2 nematodes [97]. Another study examined the impact of CBD on ROS production in mitochondria and found that CBD exposure (1 µM) decreased the human Aβ peptide-induced increase in mitochondrial ROS in the transgenic GRU102 strain [111]. Further, CBD reduced elevated ROS production in the α-synuclein and 6-OHDA nematode models of PD. Moreover, CBD enhanced the expression of the antioxidant enzyme SOD-3 and extended the lifespan of nematodes [96]. CBDV, the propyl analogue of CBD, displayed antioxidant properties by prolonging survival in the juglone-induced oxidative stress model in various nematode strains [98]. In addition, CBDV upregulated the mRNA expression of lys-7, mtl-1, sod-3, sod-4 and sod-5, which are antioxidative factors controlled by daf-16, a key transcription factor that regulates oxidative stress [98].
Structural analogues of CBD were shown to reduce ROS [97]. However, CBD-M1 (a CBD derivative with one phenolic hydroxyl group being methylated) and CBD-M2 (a CBD derivative with two methylated phenolic hydroxyl groups) were less effective than CBD itself in reducing ROS. Arguing for CBD’s antioxidant effect being due to direct scavenging of free radicals, CBD did not upregulate the expression of antioxidative genes such as catalase (CAT), superoxide dismutase (SOD) or glutathione S-transferase (GST). The synthetic analogues were also less effective than CBD in protecting against oxidative stress as indexed by juglone-induced mortality in C. elegans [97]. An interesting study administered medicinal cannabis oils to N2 worms and found antioxidant effects through a reduction in ROS, although these compounds contained a mixture of CBD and THC, and likely other compounds [112]. They did not test individual compounds alone which would have helped to deconvolute the effects observed.
In conclusion, CBD has the potential to increase the lifespan of C. elegans by promoting autophagy, decreasing oxidative stress and regulating the glyoxalase pathway. This effect is observed in both wild-type and transgenic strains, which are used to simulate neurodegenerative disorders such as AD and PD.
Exogenous plant cannabinoid effects on healthspan in C. elegans
Healthspan refers to the period of an organism’s life during which it maintains good health and functional abilities, free from age-related diseases and disorders. In short, it refers to the number of “healthy life years” [113]. In ageing research, healthspan is considered to be a more relevant and useful measure than lifespan alone, as it allows researchers to assess the impact of various interventions on the overall health and function of the organism, rather than just focusing on its lifespan. There is no point to living longer without corresponding improvements in healthy function and well-being. In C. elegans, healthspan can be assessed by measuring various parameters such as mobility, muscle function/integrity, cellular accumulation of autofluorescent pigments that reflect ageing, and resistance to stress and disease. Many studies have shown that CBD diminishes age-related decline in physiological and behavioural function in C.elegans.
As C. elegans age, they exhibit decreased locomotion due to a decline in the function of the organism’s nervous system [70], as well as a decline in muscle function [114] and other physiological processes [115]. With age, the frequency of body movements, specifically body bends, decreases in C. elegans. CBD exposure (1 µM) increased the frequency of body bends over 11 days, with a significant increase observed on day 7 [94]. Exposing C. elegans to a CBD-rich cannabis extract (MGC1013) enhanced movement patterns [116]. Despite the presence of CBD in MGC1013, the increased motility observed could not be solely attributed to CBD concentrations in the extract, as treatment with extracts derived from other CBD-rich varieties (MGC1074 and MGC1122) did not affect body bends. Furthermore, treatment with pure CBD at the same concentration as found in MGC1013 did not produce any effect [116]. This highlights that cannabis constituents beyond CBD might be explored for their beneficial effects on healthspan.
The age-related diminution of locomotor activity is accelerated in C. elegans models of neurodegenerative disease [117]. To evaluate the impact of CBD on healthspan in a disease context, a transgenic strain that expressed human Aβ1-42 in muscle tissues (CL4176) was used as an in vivo model for AD [97]. In the study, CBD delayed the paralysis of CL4176 worms in a concentration-dependent manner within a range of 10–100 µM. Notably, treatment with 100 µM CBD robustly increased body bends [97]. In another study, CBD-dominant medicinal cannabis extracts increased body bend rate compared to controls in the GMC101 nematode line where Aβ is expressed in body wall muscle [112]. This effect was also observed in heat-stressed GMC101 nematodes [112]. Further, CBD 5 µM restored exploratory behaviour deficits in pan-neuronal Aβ1-42 expressing C. elegans [95]. CBD (25, 50 and 100 µM) concentration-dependently improved deficits in food-sensing and locomotory rate in the 6-OHDA model of PD in C. elegans [96]. Further, CBDV (50 µM) mitigated the impaired body bending observed in another PD model strain (NL5901) that expresses human α-synuclein protein in body wall muscle [98]. Notably, CBDV did not delay the onset of paralysis in an AD nematode model at the same concentration.
Pharyngeal pumping is a well-characterized feeding behaviour in C. elegans and is considered a healthspan indicator that reflects the organism’s ability to ingest and process nutrients [118]. The pharyngeal pumping rate declines with age in C. elegans [119]. Studies have demonstrated that CBD increased the pumping rate in normal ageing and age-related diseases. In normal ageing, 1 µM CBD increased the pharyngeal pumping rate on days 3 and 5 of repeated daily treatment [94]. Similar results were observed when testing CBD-dominant cannabis extracts in the N2 worm strain [112]. In CBD-rich varieties, only one chemotype (MGC 1013) increased the pumping rate in a dose-dependent manner [116]. It is noteworthy that a polar fraction of the extract was responsible for the effects, suggesting a role for other non-cannabinoid constituents, as the cannabinoids are non-polar [116]. The effects of CBD on pharyngeal pumping rate have also been examined in AD models. Treatment with both low (1 µM and 5 µM) [95, 111] and high (100 µM) [97] CBD concentrations enhanced pumping rate in models where Aβ was overexpressed in muscle and neurons. In a PD model that overexpresses α-synuclein in muscle (NL5901), CBDV 50 µM diminished the age-related decline in pumping rate [98]. The reproductive system of C. elegans shows a decrease during ageing [120]. Various approaches have been discovered that slow reproductive ageing, such as genetic mutations [121], small molecules [122] and environmental factors [123]. CBD exposure (1 µM and 5 µM) increased the number of laid eggs and increased the number of progeny (brood size) compared to the control group in both wild-type and Aβ transgenic worms [94, 95].
There is emerging evidence that CBD has neuroprotective effects in C. elegans. The normal ageing of neurons in C. elegans involves subtle morphological changes with no apparent neuron loss or axon deficit [124]. These morphological changes in neurons include irregularly shaped somas, disorganized microtubule arrangements and the occurrence of bubble-like lesions, beading and blebs resulting in the distortion of the neuron structure [69, 124]. These changes have been found in touch receptor neurons (TRNs), as well as dopaminergic neurons [69, 124]. A recent study investigated the impact of CBD on the normal ageing of both anterior (ALM) and posterior (PLM) TRN neurons in a transgenic strain expressing a TRN-specific mec-4p::GFP [94]. CBD treatment reduced the number of irregularly shaped somas of ALM neurons in both young and old worms. However, CBD only reduced the proportion of defective processes (branching, beading and blebbing) in PLM neurons in aged worms but not young worms. In a disease context, CBD derivatives with methylated phenolic hydroxyl groups (CBD-M1 and CBD-M2) mitigated Aβ-induced neurotoxicity, showcasing CBD’s remarkable neuroprotective effects [95, 97]. Further, CBD also protected dopaminergic neurons from reserpine-induced degeneration and locomotor deficits, which were associated with reductions in ROS and α-syn accumulation [125].
Studies have also addressed the impact of cannabis and cannabinoids on sensory and neurobehavioural function. A study found that cannabis extracts had varying effects on sensory responses and memory in worms [116]. The cannabis extracts reduced the response to a noxious odour, which if anything suggests a reduction in sensory function. However, a CBD-rich variety extract also reduced adverse memory responses in nematodes [116]. Neuronal Aβ expression induces toxicity, leading C. elegans to exhibit deficiencies in their chemotaxis behavior [126]. CBD at both low (5 µM) and high (100 µM) concentrations increased chemotaxis indexes in Aβ overexpressed strains [95, 97].
Anti-ageing effects of cannabinoids in fruit fly, zebrafish and mice
A recent study assessed the impact of cannabinoids on ageing in Drosophila. Flies treated with 3 µM CBD had significantly increased lifespan, whereas THC at the same concentration resulted in a modest decrease in longevity [127] (Fig. 6). Although CBD or THC at various concentrations had little effect on sleep and circadian-based behaviors, and the age-dependent decline in locomotor activity. Cannabinoids have been assessed for their effects on lifespan and healthspan in zebrafish (Fig. 6). THC increased survival of aged (30 months of age) male zebrafish at a low concentration (80 nM) but not at higher concentrations (400 nM and 2 µM) [128]. The low concentration of THC reduced body weight, spinal deformity and the liver expression of inflammatory and senescence markers (p16ink4ab, TNF-α, IL-1β, IL-6, PPARα and PPARγ). Moreover, it increased egg production, while the higher THC concentrations impaired fecundity. Another study showed that exposure to CBD (0.02, 0.1, 0.5 μM) during the F0 generation in zebrafish led to a remarkable increase in survival. Concentrations of 100 and 500 nM CBD increased survival in male zebrafish, but not female zebrafish. Although there was a strong non-significant trend of increased survival in the females with as great as a 50% increase in survival at 100 nM; it is possible that the study was underpowered. Although the 500 nM CBD concentration reduced sperm count in the males, 100 nM CBD increased egg production in the female fish. Additionally, CBD reduced the weight and length of zebrafish, although CBD reduced age-related increases in the inflammatory markers TNF-α and IL-1β in males and females. Unlike THC, there was no effect on the senescence-associated markers p16 or IL-6.

An overview of the anti-aging effects of CBD and THC in rodents, zebrafish, C. elegans and Drosophila
The available data on the effect of cannabinoids in rodent longevity is very limited. The study that assessed the impact of THC on rodent lifespan was conducted by the US National Toxicology Program (NTP) [129, 130]. In this study, mice and rats were dosed with relatively high oral doses of THC (12.5 mg/kg, 25 mg/kg and 50 mg/kg) over 2 years. THC considerably increased the lifespan of both male and female rats (Fig. 7). Although, no survival benefit of THC was observed in the mouse study. To the best of our knowledge, the effect of CBD on lifespan in rodents has not been examined. Although, there is evidence that CBD might improve healthspan in rodents. For example, one interesting study conducted on “middle-aged” rats (15 months old) showed that long-term treatment with CBD had beneficial effects on lung and liver health by lowering age-related oxidative stress and pro-apoptotic and inflammatory markers [131] (Fig. 6). Repeated CBD administration also had beneficial effects on emotional function and brain health in middle-aged female rats, where CBD reduced social isolation-induced behavioural despair and loss of brain-derived neurotrophic factor (BDNF) in the ventral striatum [132]. Consistent with this finding, repeated CBD treatment reduced behaviour despair in the forced-swim test in “older” rats (19–21 months of age) [133]. In addition, repeated CBD treatment improved blood glucose concentrations, nullified memory impairment and normalised neurodegeneration and neuroinflammation in response to chronic cerebral hypofusion in middle-aged diabetic rats [134]. It should also be pointed out that CBD and THC exposure promoted cognitive enhancing effects and improved brain function in mouse models of Alzheimer’s disease (for details see excellent reviews on this topic [135,136,137]).

The effects of cannabinoids on mechanisms of ageing
There is an intense research effort in geroscience devoted to characterising the cellular and molecular mechanisms of ageing. The hallmarks of ageing include stem cell exhaustion, loss of proteostasis, mitochondrial dysfunction, genome instability, telomere attrition, epigenetic changes and cellular senescence [138]. Future studies are needed to assess whether cannabinoids can revert the hallmarks of ageing. We are beginning to see research in this area, with recent studies assessing whether cannabinoids affect cellular senescence. Preclinical research has shown that genetic and pharmacological deletion of senescent cells can extend lifespan and healthspan in animal studies [138]. Senescent cells lose the ability to divide due to telomere attrition and release mediators associated with the senescence-associated secretory phenotype (SASP) which promotes inflammation and cell damage in neighbouring healthy tissues. Thus, there is interest in developing agents that have senolytic (the ability to kill senescence cells) or senomorphic (the ability to diminish the SASP) activity.
In one study, both THC and CBD were examined in healthy and stress-induced premature senescent skin fibroblasts yielding perplexing results, with the title of the paper stating the cannabinoids prevented cellular senescence, but the balance of the data in the paper showing that the cannabinoids increased the viability of senescent cells [139]. Under conditions of replicative senescence, a cellular model of healthy ageing, treatment with THC (0.25–10 µM) non-dose-dependently increased senescence-associated β-galactosidase (SA-β-gal) staining following repeated, daily 2 h incubations for 2 and 15 days. Interestingly, CBD also non-dose dependently increased replicative senescent cell number as gauged by SA-β-gal after 2 days of treatment, although following 15 days of treatment CBD 10 µM reduced SA-β-gal activity. The authors themselves questioned whether this was a beneficial effect, and stated it was likely explained by CBD having overall cytotoxic effects on the cells, thus non-specifically decreasing all cell populations. CBD and THC also increased the viability of the skin fibroblasts that were induced to senescence with hydrogen peroxide; again, this might suggest that the cannabinoids increase the viability of senescent cells. However, 5 days of treatment with 2 µM CBD and THC reduced SA-β-gal in hydrogen peroxide-induced senescent cells. CBD and THC also enhanced wound healing of healthy and senescent skin fibroblast cells in the scratch assay. Notably, THC and CBD showed remarkable effectiveness in enhancing wound healing when compared to well-known anti-ageing nutrient signaling regulators like metformin, rapamycin and triacetylresveratrol.
In a follow-up study by the same lab, it was shown that the combination of CBD with triacetylresveratrol yielded significant benefits in senescent fibroblasts, by increasing viability and improving wound healing [140]. Notably, these studies also presented data highlighting that THC and CBD might have sirtuin-activating effects. SIRTs are signaling proteins that, among many functions, repair age-related DNA damage, and their activation has been suggested to extend lifespan and healthspan in model systems, albeit with some controversy [141, 142]. THC increased SIRT6 and CBD, when combined with triacetylresveratrol increased SIRT1 mRNA in skin fibroblasts [139, 140].
CBD treatment triggered cellular senescence in primary human Sertoli cells, a condition associated with male reproductive toxicity [143]. This effect was evident as CBD enhanced the expression of SA-β-gal and upregulated a set of genes related to SASP, along with activating the p53 signaling pathway [143]. In human astrocytes, CBD protected against Aβ-induced mitochondrial ROS and cellular senescence as gauged by SA-β-gal staining and senescence markers p16, p21 and p53 [111]. These protective effects of CBD were attributed to the activation of Parkin-dependent mitophagy which is autophagy specific to the removal of damaged mitochondria. In conclusion, these results highlight that cannabinoids do influence cellular senescence, but more research is needed to further illuminate the nature of the relationship. It would also be of interest to assess the effects of the cannabinoids on other hallmarks of ageing.
Conclusion
Ageing is a complex and multifactorial process that occurs as a gradual accumulation of cellular damage in various tissues of the body, leading to a decline in physiological functions across all systems. One main aim of ageing research is to identify compounds that can postpone deteriorative changes linked to ageing. Finding interventions that promote healthy ageing may provide a paradigm shift in medicine, by targeting the common denominator of many diseases, that is, the ageing process. With the ongoing trend of cannabis legalisation globally, there is a demand for research that explores the impact of cannabis and cannabinoids on healthy ageing and diseases of ageing. The current review highlights that cannabinoids, whether endogenous or exogenous, extend lifespan and healthspan in model systems. However, more research is needed to observe whether these results translate in mammalian systems and ultimately in the clinic. The anti-ageing effects of cannabinoids have a number of different mechanisms, including the reduction of oxidative stress and the triggering of autophagy. More research is needed to further explore the anti-ageing mechanisms of the cannabinoids and to more comprehensively examine their impact on the hallmarks of ageing including cellular senescence. The development of anti-ageing agents that tone up endocannabinoid transmission could also be examined. Moreover, plant cannabinoids beyond CBD could be explored, as well as other cannabis constituents, alone and in combination. In addition, given the robust findings with CBD, chemical analogues of CBD might be developed. The current review underscores that there is much promise for the further development of cannabinoids as anti-ageing agents, to improve healthy ageing and general well-being.
Data Availability
All the data is available on the request of the authors.
References
Kaeberlein M. Longevity and aging. F1000Prime Rep. 2013;5:5.
Lopez-Otin C, et al. The hallmarks of aging. Cell. 2013;153(6):1194–217.
North BJ, Sinclair DA. The intersection between aging and cardiovascular disease. Circ Res. 2012;110(8):1097–108.
Ahima RS. Connecting obesity, aging and diabetes. Nat Med. 2009;15(9):996–7.
Hoeijmakers JH. DNA damage, aging, and cancer. N Engl J Med. 2009;361(15):1475–85.
Franceschi C, et al. Inflammaging and “Garb-aging.” Trends Endocrinol Metab. 2017;28(3):199–212.
Paneni F, et al. The aging cardiovascular system: understanding it at the cellular and clinical levels. J Am Coll Cardiol. 2017;69(15):1952–67.
Alcedo J, Flatt T, Pasyukova EG. The role of the nervous system in aging and longevity. Front Genet. 2013;4:124.
Murman DL. The impact of age on cognition. Semin Hear. 2015;36(3):111–21.
Cavazzana A, et al. Sensory-specific impairment among older people. An investigation using both sensory thresholds and subjective measures across the five senses. PLoS One. 2018; 13(8):e0202969.
Arkell TR, et al. Assessment of medical cannabis and health-related quality of life. JAMA Netw Open. 2023;6(5):e2312522.
Abyadeh M, et al. A proteomic view of cellular and molecular effects of cannabis. Biomolecules. 2021;11(10):1411
Arnold JC. A primer on medicinal cannabis safety and potential adverse effects. Aust J Gen Pract. 2021;50(6):345–50.
Mulligan MK, Hamre KM. Influence of prenatal cannabinoid exposure on early development and beyond. Adv Drug Alcohol Res. 2023;3:10981.
Scheyer AF, et al. Cannabis in adolescence: lasting cognitive alterations and underlying mechanisms. Cannabis Cannabinoid Res. 2023;8(1):12–23.
Pertwee RG. Cannabinoid pharmacology: the first 66 years. Br J Pharmacol. 2006;147:S163–71.
Banister SD, et al. Dark classics in chemical neuroscience: delta(9)-tetrahydrocannabinol. ACS Chem Neurosci. 2019;10(5):2160–75.
Arnold JC, et al. The safety and efficacy of low oral doses of cannabidiol: an evaluation of the evidence. Clin Transl Sci. 2023;16(1):10–30.
Arnold JC, Nation T, McGregor IS. Prescribing medicinal cannabis. Aust Prescr. 2020;43(5):152–9.
Wise J. FDA approves its first cannabis based medicine. BMJ. 2018;361: k2827.
Wise J. European drug agency approves cannabis-based medicine for severe forms of epilepsy. BMJ. 2019;366:l5708.
Anderson LL, et al. Pharmacokinetics of phytocannabinoid acids and anticonvulsant effect of cannabidiolic acid in a mouse model of Dravet syndrome. J Nat Prod. 2019;82(11):3047–55.
Anderson LL, et al. Cannabichromene, related phytocannabinoids, and 5-fluoro-cannabichromene have anticonvulsant properties in a mouse model of Dravet syndrome. ACS Chem Neurosci. 2021;12(2):330–9.
Anderson LL, et al. Cannabigerolic acid, a major biosynthetic precursor molecule in cannabis, exhibits divergent effects on seizures in mouse models of epilepsy. Br J Pharmacol. 2021;178(24):4826–41.
Assareh N, et al. Cannabidiol disrupts conditioned fear expression and cannabidiolic acid reduces trauma-induced anxiety-related behaviour in mice. Behav Pharmacol. 2020;31(6):591–6.
Hill AJ, et al. Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacol Ther. 2012;133(1):79–97.
Di Marzo V, Piscitelli F. The endocannabinoid system and its modulation by phytocannabinoids. Neurotherapeutics. 2015;12(4):692–8.
Kesner AJ, Lovinger DM. Cannabinoids, endocannabinoids and sleep. Front Mol Neurosci. 2020;13:125.
Rakotoarivelo V, Sihag J, Flamand N. Role of the endocannabinoid system in the adipose tissue with focus on energy metabolism. Cells. 2021;10(6):1279.
Ashton CH, Moore PB. Endocannabinoid system dysfunction in mood and related disorders. Acta Psychiatr Scand. 2011;124(4):250–61.
Almogi-Hazan O, Or R. Cannabis, the endocannabinoid system and immunity-the journey from the bedside to the bench and back. Int J Mol Sci. 2020;21(12):4448.
Pacher P, Batkai S, Kunos G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58(3):389–462.
Zou S, Kumar U. Cannabinoid receptors and the endocannabinoid system: signaling and function in the central nervous system. Int J Mol Sci. 2018;19(3):833.
Elmes MW, et al. Fatty acid-binding proteins (FABPs) are intracellular carriers for Delta9-tetrahydrocannabinol (THC) and cannabidiol (CBD). J Biol Chem. 2015;290(14):8711–21.
Berger WT, et al. Targeting fatty acid binding protein (FABP) anandamide transporters—a novel strategy for development of anti-inflammatory and anti-nociceptive drugs. PLoS ONE. 2012;7(12):e50968.
Haj-Dahmane S, et al. Fatty-acid-binding protein 5 controls retrograde endocannabinoid signaling at central glutamate synapses. Proc Natl Acad Sci U S A. 2018;115(13):3482–7.
Anderson LL, et al. Coadministered cannabidiol and clobazam: preclinical evidence for both pharmacodynamic and pharmacokinetic interactions. Epilepsia. 2019;60(11):2224–34.
Bakas T, et al. The direct actions of cannabidiol and 2-arachidonoyl glycerol at GABA(A) receptors. Pharmacol Res. 2017;119:358–70.
Anavi-Goffer S, et al. Modulation of L-alpha-lysophosphatidylinositol/GPR55 mitogen-activated protein kinase (MAPK) signaling by cannabinoids. J Biol Chem. 2012;287(1):91–104.
Gray RA, et al. Anticonvulsive properties of cannabidiol in a model of generalized seizure are transient receptor potential vanilloid 1 dependent. Cannabis Cannabinoid Res. 2020;5(2):145–9.
Gray RA, Whalley BJ. The proposed mechanisms of action of CBD in epilepsy. Epileptic Disord. 2020;22(S1):10–5.
Kaplan JS, et al. Cannabidiol attenuates seizures and social deficits in a mouse model of Dravet syndrome. Proc Natl Acad Sci U S A. 2017;114(42):11229–34.
Choi K, et al. Expression pattern of the cannabinoid receptor genes in the frontal cortex of mood disorder patients and mice selectively bred for high and low fear. J Psychiatr Res. 2012;46(7):882–9.
Berrendero F, et al. Changes in cannabinoid receptor binding and mRNA levels in several brain regions of aged rats. Biochim Biophys Acta. 1998;1407(3):205–14.
Li X, et al. Lifespan changes in cannabinoid 1 receptor mRNA expression in the female C57BL/6J mouse brain. J Comp Neurol. 2023;531(2):294–313.
Bilkei-Gorzo A, et al. Early age-related cognitive impairment in mice lacking cannabinoid CB1 receptors. Proc Natl Acad Sci U S A. 2005;102(43):15670–5.
Nidadavolu P, et al. Dynamic changes in the endocannabinoid system during the aging process: focus on the middle-age crisis. Int J Mol Sci. 2022;23(18):10254.
Albayram O, Bilkei-Gorzo A, Zimmer A. Loss of CB1 receptors leads to differential age-related changes in reward-driven learning and memory. Front Aging Neurosci. 2012;4:34.
Wang L, et al. Endocannabinoid signaling via cannabinoid receptor 1 is involved in ethanol preference and its age-dependent decline in mice. Proc Natl Acad Sci U S A. 2003;100(3):1393–8.
Komorowska-Muller JA, et al. Cannabinoid receptor 2 alters social memory and microglial activity in an age-dependent manner. Molecules. 2021;26(19):5984.
Lindsey LP, et al. The cannabinoid receptor 2 agonist, beta-caryophyllene, improves working memory and reduces circulating levels of specific proinflammatory cytokines in aged male mice. Behav Brain Res. 2019;372: 112012.
Long LE, et al. Developmental trajectory of the endocannabinoid system in human dorsolateral prefrontal cortex. BMC Neurosci. 2012;13:87.
Rossi G, et al. Endocannabinoid system components of the female mouse reproductive tract are modulated during reproductive aging. Int J Mol Sci. 2023;24(8):7542.
He Y, Jasper H. Studying aging in Drosophila. Methods. 2014;68(1):129–33.
Piper MDW, Partridge L. Drosophila as a model for ageing. Biochim Biophys Acta Mol Basis Dis. 2018;1864(9 Pt A):2707–17.
Olsen A, Vantipalli MC, Lithgow GJ. Using Caenorhabditis elegans as a model for aging and age-related diseases. Ann N Y Acad Sci. 2006;1067:120–8.
Van Houcke J, et al. The zebrafish as a gerontology model in nervous system aging, disease, and repair. Ageing Res Rev. 2015;24(Pt B):358–68.
Choi TY, et al. Zebrafish as an animal model for biomedical research. Exp Mol Med. 2021;53(3):310–7.
Zhang S, et al. Caenorhabditis elegans as a useful model for studying aging mutations. Front Endocrinol (Lausanne). 2020;11:554994.
Wangler MF, Yamamoto S, Bellen HJ. Fruit flies in biomedical research. Genetics. 2015;199(3):639–53.
Folgueras AR, et al. Mouse models to disentangle the hallmarks of human aging. Circ Res. 2018;123(7):905–24.
Crimmins EM. Lifespan and healthspan: past, present, and promise. Gerontologist. 2015;55(6):901–11.
Son HG, et al. Age-dependent changes and biomarkers of aging in Caenorhabditis elegans. Aging Cell. 2019;18(2):e12853.
Herndon LA, et al. Effects of ageing on the basic biology and anatomy of C. elegans. ageing: lessons from C. elegans. Springer International Publishing: Cham, Switzerland. 2017; p. 9–39.
Andux S, Ellis RE. Apoptosis maintains oocyte quality in aging Caenorhabditis elegans females. PLoS Genet. 2008;4(12):e1000295.
Hughes SE, et al. Genetic and pharmacological factors that influence reproductive aging in nematodes. PLoS Genet. 2007;3(2): e25.
Chen J, et al. A demographic analysis of the fitness cost of extended longevity in Caenorhabditis elegans. J Gerontol A Biol Sci Med Sci. 2007;62(2):126–35.
Lee Y, et al. Inverse correlation between longevity and developmental rate among wild C. elegans strains. Aging. 2016;8(5):986–99.
Chew YL, et al. Aging in the nervous system of Caenorhabditis elegans. Commun Integr Biol. 2013;6(5):e25288.
Toth ML, et al. Neurite sprouting and synapse deterioration in the aging Caenorhabditis elegans nervous system. J Neurosci. 2012;32(26):8778–90.
De Haes W, et al. Metformin promotes lifespan through mitohormesis via the peroxiredoxin PRDX-2. Proc Natl Acad Sci U S A. 2014;111(24):E2501–9.
Xenos D, Mecocci P, Boccardi V. A blast from the past: to tame time with metformin. Mech Ageing Dev. 2022;208:111743.
Zhang K, et al. Metformin and risk of cancer among patients with type 2 diabetes mellitus: a systematic review and meta-analysis. Prim Care Diabetes. 2021;15(1):52–8.
Campbell JM, et al. Metformin use associated with reduced risk of dementia in patients with diabetes: a systematic review and meta-analysis. J Alzheimers Dis. 2018;65(4):1225–36.
Johnson JA, et al. Reduced cardiovascular morbidity and mortality associated with metformin use in subjects with Type 2 diabetes. Diabet Med. 2005;22(4):497–502.
Kulkarni AS, et al. Metformin regulates metabolic and nonmetabolic pathways in skeletal muscle and subcutaneous adipose tissues of older adults. Aging Cell. 2018;17(2):e12723.
Lehtonen M, et al. Mass-spectrometric identification of anandamide and 2-arachidonoylglycerol in nematodes. Chem Biodivers. 2008;5(11):2431–41.
Basavarajappa BS. Critical enzymes involved in endocannabinoid metabolism. Protein Pept Lett. 2007;14(3):237–46.
Lucanic M, et al. N-acylethanolamine signalling mediates the effect of diet on lifespan in Caenorhabditis elegans. Nature. 2011;473(7346):226–9.
Harrison N, et al. Characterization of N-acyl phosphatidylethanolamine-specific phospholipase-D isoforms in the nematode Caenorhabditis elegans. PLoS ONE. 2014;9(11):e113007.
Lakowski B, Hekimi S. The genetics of caloric restriction in Caenorhabditis elegans. Proc Natl Acad Sci U S A. 1998;95(22):13091–6.
Bishop NA, Guarente L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat Rev Genet. 2007;8(11):835–44.
Lin YH, et al. Diacylglycerol lipase regulates lifespan and oxidative stress response by inversely modulating TOR signaling in Drosophila and C. elegans. Aging Cell. 2014;13(4):755–64.
Chen AL, et al. Pharmacological convergence reveals a lipid pathway that regulates C. elegans lifespan. Nat Chem Biol. 2019;15(5):453–62.
Pastuhov SI, Matsumoto K, Hisamoto N. Endocannabinoid signaling regulates regenerative axon navigation in Caenorhabditis elegans via the GPCRs NPR-19 and NPR-32. Genes Cells. 2016;21(7):696–705.
Crooks BA, et al. Pan-phylum in silico analyses of nematode endocannabinoid signalling systems highlight novel opportunities for parasite drug target discovery. Front Endocrinol (Lausanne). 2022;13:892758.
Oakes MD, et al. Cannabinoids activate monoaminergic signaling to modulate key C. elegans behaviors. J Neurosci. 2017;37(11):2859–69.
Hernandez-Cravero B, et al. Cannabinoids activate the insulin pathway to modulate mobilization of cholesterol in C. elegans. PLoS Genet. 2022;18(11):e1010346.
Galles C, et al. Endocannabinoids in Caenorhabditis elegans are essential for the mobilization of cholesterol from internal reserves. Sci Rep. 2018;8(1):6398.
Levichev A, et al. The conserved endocannabinoid anandamide modulates olfactory sensitivity to induce hedonic feeding in C. elegans. Curr Biol. 2023;33(9):1625–39.e4.
McGregor IS, et al. Access to cannabidiol without a prescription: a cross-country comparison and analysis. Int J Drug Policy. 2020;85:102935.
Land MH, et al. Effect of cannabidiol on the long-term toxicity and lifespan in the preclinical model Caenorhabditis elegans. Cannabis Cannabinoid Res. 2021;6(6):522–7.
Frandsen J, Narayanasamy P. Effect of cannabidiol on the neural glyoxalase pathway function and longevity of several C. elegans strains including a C. elegans Alzheimer’s disease model. ACS Chem Neurosci. 2022;13(8):1165–77.
Wang Z, et al. Cannabidiol induces autophagy and improves neuronal health associated with SIRT1 mediated longevity. Geroscience. 2022;44(3):1505–24.
Wang Z, et al. Cannabidiol regulates CB1-pSTAT3 signaling for neurite outgrowth, prolongs lifespan, and improves health span in Caenorhabditis elegans of Abeta pathology models. FASEB J. 2021;35(5):e21537.
Muhammad F, et al. Neuroprotective effects of cannabidiol on dopaminergic neurodegeneration and alpha-synuclein accumulation in C. elegans models of Parkinson’s disease. Neurotoxicology. 2022;93:128–39.
Zhang Y, et al. Cannabidiol protects against Alzheimer’s disease in C. elegans via ROS scavenging activity of its phenolic hydroxyl groups. Eur J Pharmacol. 2022;919:174829.
Wang F, et al. Cannabidivarin alleviates alpha-synuclein aggregation via DAF-16 in Caenorhabditis elegans. FASEB J. 2023;37(2):e22735.
Lucanic M, et al. Impact of genetic background and experimental reproducibility on identifying chemical compounds with robust longevity effects. Nat Commun. 2017;8:14256.
Hou Y, et al. Ageing as a risk factor for neurodegenerative disease. Nat Rev Neurol. 2019;15(10):565–81.
Nobili A, et al. Dopamine neuronal loss contributes to memory and reward dysfunction in a model of Alzheimer’s disease. Nat Commun. 2017;8:14727.
Mamelak M. Parkinson’s disease, the dopaminergic neuron and gammahydroxybutyrate. Neurol Ther. 2018;7(1):5–11.
Hansen M, Rubinsztein DC, Walker DW. Autophagy as a promoter of longevity: insights from model organisms. Nat Rev Mol Cell Biol. 2018;19(9):579–93.
Nakamura S, Yoshimori T. Autophagy and Longevity. Mol Cells. 2018;41(1):65–72.
Aman Y, et al. Autophagy in healthy aging and disease. Nat Aging. 2021;1(8):634–50.
Chen Y, Scarcelli V, Legouis R. Approaches for studying autophagy in Caenorhabditis elegans. Cells. 2017; 6(3):27.
Melendez A, Levine B. Autophagy in C. elegans. WormBook. 2009; 1–26.
Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014;24(10):R453–62.
Davalli P, et al. ROS, Cell senescence, and novel molecular mechanisms in aging and age-related diseases. Oxid Med Cell Longev. 2016;2016:3565127.
Aragones G, et al. The glyoxalase system in age-related diseases: nutritional intervention as anti-ageing strategy. Cells. 2021; 10(8):1852.
Wang Z, et al. Parkin mediates cannabidiol prevention of amyloid-beta-induced senescence in human astrocytes. Cannabis Cannabinoid Res. 2023;8(2):309–20.
Vanin AP, et al. Neuroprotective potential of Cannabis sativa-based oils in Caenorhabditis elegans. Sci Rep. 2022;12(1):15376.
Crane PA, Wilkinson G, Teare H. Healthspan versus lifespan: new medicines to close the gap. Nat Aging. 2022;2(11):984–8.
Guo X, et al. Behavioral decay in aging male C. elegans correlates with increased cell excitability. Neurobiol Aging. 2012;33(7):1483e5-1523.
Glenn CF, et al. Behavioral deficits during early stages of aging in Caenorhabditis elegans result from locomotory deficits possibly linked to muscle frailty. J Gerontol A Biol Sci Med Sci. 2004;59(12):1251–60.
van Es-Remers M, et al. C. elegans as a test system to study relevant compounds that contribute to the specific health-related effects of different cannabis varieties. J Cannabis Res. 2022;4(1):53.
Alexander AG, Marfil V, Li C. Use of Caenorhabditis elegans as a model to study Alzheimer’s disease and other neurodegenerative diseases. Front Genet. 2014;5:279.
Raizen D, et al. Methods for measuring pharyngeal behaviors. WormBook. 2012;1–13.
Huang C, Xiong C, Kornfeld K. Measurements of age-related changes of physiological processes that predict lifespan of Caenorhabditis elegans. Proc Natl Acad Sci U S A. 2004;101(21):8084–9.
Scharf A, et al. Reproductive aging in caenorhabditis elegans: from molecules to ecology. Front Cell Dev Biol. 2021;9:718522.
Hughes SE, Huang C, Kornfeld K. Identification of mutations that delay somatic or reproductive aging of Caenorhabditis elegans. Genetics. 2011;189(1):341–56.
Ryu D, et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat Med. 2016;22(8):879–88.
Huang TT, et al. Age-dependent changes in response property and morphology of a thermosensory neuron and thermotaxis behavior in Caenorhabditis elegans. Aging Cell. 2020;19(5):e13146.
Pan CL, et al. Genetic analysis of age-dependent defects of the Caenorhabditis elegans touch receptor neurons. Proc Natl Acad Sci U S A. 2011;108(22):9274–9.
da Cruz Guedes E, et al. Cannabidiol recovers dopaminergic neuronal damage induced by reserpine or alpha-synuclein in Caenorhabditis elegans. Neurochem Res. 2023;48(8):2390–405.
Wu Y, et al. Amyloid-beta-induced pathological behaviors are suppressed by Ginkgo biloba extract EGb 761 and ginkgolides in transgenic Caenorhabditis elegans. J Neurosci. 2006;26(50):13102–13.
Candib A, et al. The influence of cannabinoids on drosophila behaviors, longevity, and traumatic injury responses of the adult nervous system. Cannabis Cannabinoid Res. 2023.
Pandelides Z, et al. Developmental exposure to Delta(9)-tetrahydrocannabinol (THC) causes biphasic effects on longevity, inflammation, and reproduction in aged zebrafish (Danio rerio). Geroscience. 2020;42(3):923–36.
National Toxicology P. NTP Toxicology and Carcinogenesis Studies of 1-Trans-Delta(9)-Tetrahydrocannabinol (CAS No. 1972–08–3) in F344 Rats and B6C3F1 Mice (Gavage Studies). Natl Toxicol Program Tech Rep Ser. 1996;446:1–317.
Chan PC, et al. Toxicity and carcinogenicity of delta 9-tetrahydrocannabinol in Fischer rats and B6C3F1 mice. Fundam Appl Toxicol. 1996;30(1):109–17.
Rancan L, et al. Protective actions of cannabidiol on aging-related inflammation, oxidative stress and apoptosis alterations in liver and lung of long evans rats. Antioxidants (Basel). 2023;12(10):1837.
Saad N, et al. Cannabidiol modulates emotional function and brain-derived neurotrophic factor expression in middle-aged female rats exposed to social isolation. Int J Mol Sci. 2023;24(20):15492.
Hernandez-Hernandez E, Garcia-Fuster MJ. Dose-dependent antidepressant-like effects of cannabidiol in aged rats. Front Pharmacol. 2022;13:891842.
Santiago AN, et al. Effects of cannabidiol on diabetes outcomes and chronic cerebral hypoperfusion comorbidities in middle-aged rats. Neurotox Res. 2019;35(2):463–74.
Karl T, et al. The therapeutic potential of the endocannabinoid system for Alzheimer’s disease. Expert Opin Ther Targets. 2012;16(4):407–20.
Karl T, Garner B, Cheng D. The therapeutic potential of the phytocannabinoid cannabidiol for Alzheimer’s disease. Behav Pharmacol. 2017;28(2 and 3-Spec Issue):142–60.
Coles M, Steiner-Lim GZ, Karl T. Therapeutic properties of multi-cannabinoid treatment strategies for Alzheimer’s disease. Front Neurosci. 2022;16:962922.
Wissler Gerdes EO, et al. Strategies for late phase preclinical and early clinical trials of senolytics. Mech Ageing Dev. 2021;200:111591.
Gerasymchuk M, et al. Phytocannabinoids stimulate rejuvenation and prevent cellular senescence in human dermal fibroblasts. Cells. 2022;11(23):3939.
Gerasymchuk M, et al. The effects of nutrient signaling regulators in combination with phytocannabinoids on the senescence-associated phenotype in human dermal fibroblasts. Int J Mol Sci. 2022;23(15):8804.
Brenner C. Sirtuins are not conserved longevity genes. Life Metab. 2022;1(2):122–33.
Bonkowski MS, Sinclair DA. Slowing ageing by design: the rise of NAD(+) and sirtuin-activating compounds. Nat Rev Mol Cell Biol. 2016;17(11):679–90.
Li Y, et al. Cannabidiol-induced transcriptomic changes and cellular senescence in human Sertoli cells. Toxicol Sci. 2023;191(2):227–38.
Acknowledgements
The authors gratefully acknowledge Barry and Joy Lambert for their continued support of the Lambert Initiative for Cannabinoid Therapeutics.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This study received funding from the Lambert Initiative for Cannabinoid Therapeutics.
Author information
Authors and Affiliations
Contributions
All authors were involved in the review concept and design. ZW was responsible for literature searching and first draft formulation. JA was responsible for writing, revising and final approval of the published manuscript.
Corresponding author
Ethics declarations
Competing interests
JCA is Director of Preclinical Research at the Lambert Initiative for Cannabinoid Therapeutics, a philanthropically funded research program at the University of Sydney. He has served as an expert witness in various medicolegal cases involving cannabis and has received consulting fees from the World Health Organisation (WHO), Medical Cannabis Industry Australia (MCIA) and Haleon (consumer healthcare subsidiary of Glaxo Smith-Kline). He reports research grants from the Australian National Health and Medical Research Council (NHMRC) and from Lambert Initiative for Cannabinoid Therapeutics. He is an inventor on patents WO2019227167 and WO2019071302 issued, which relate to cannabinoid therapeutics. All other authors have no competing financial or non-financial interests to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Wang, Z., Arnold, J.C. Cannabinoids and healthy ageing: the potential for extending healthspan and lifespan in preclinical models with an emphasis on Caenorhabditis elegans. GeroScience (2024). https://doi.org/10.1007/s11357-024-01162-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11357-024-01162-8